
昆虫的气味受体蛋白在细胞膜上形成离子通道,受体与气味分子结合时通道打开。解析非激活和激活状态下通道的结构有助于深入了解昆虫如何检测和区分不同的气味。
当使用驱虫剂来阻止昆虫传播的疾病时,我们可能会思考这些物质是如何起作用的,它们怎么才能更有效。避蚊胺是驱虫剂中最常用的化合物,被认为可以广泛激活昆虫的气味受体蛋白,扰乱昆虫寻找宿主的嗅觉编码[1]。但尚不清楚避蚊胺或天然气味分子如何结合并影响昆虫气味受体的活性。在《自然》发表的一篇文章中,del Mármol 等人[2]报道了与避蚊胺或丁香酚相关的昆虫气味受体的结构,为气味如何与受体结合,以及激活受体的结构随后如何变化提供了关键见解。
不同动物的嗅觉系统已经进化到可以完成高度专业化的任务。果蝇专精腐烂的水果,蚊子寻找宿主,而人类可以区分大量与食物有关的气味。脊椎动物和无脊椎动物都使用一组众多气味受体来检测和区分数量庞大的挥发性化学物质。许多情况下,受体-气味的结合是混杂的:也就是说,单一气味可能激活多个受体,而每个受体可能被多个气味激活。嗅觉神经元细胞各表达一种气味受体。因此,每种气味都可以激活一组不同的(但有时是重叠的)神经元,从而产生一个组合编码,供神经系统破译[3-5]。
果蝇和人类气味编码的基本原则非常相似。例如在这两个物种中,表达相同受体的嗅觉神经元的投射会聚在称为嗅小球的枢纽状结构处。然而,昆虫和脊椎动物中的受体蛋白非常不同。脊椎动物气味受体是 G 蛋白偶联受体家族的成员[6],而昆虫的受体是一些与气味分子结合时会打开的离子通道[7-10]。
几十年来,这一现象人们早已经知晓,但尚不清楚单个气味受体如何对如此数量庞大结构不同的分子做出反应。1894年Emil Fischer首次提出受体-配体结合的锁钥模型,该模型假定配体分子的形状及其在受体中的结合位点完全互补[11]。但这不足以解释受体的混杂性[12]。要揭示这一现象,需要了解与不同气味配体结合的受体的结构。
为了实现这一目标,del Mármol等人专注于昆虫气味受体。大多数此类受体由不同亚基(即异源多聚体)组装而成,所有受体均包含一个特定亚基Orco和另一个赋予配体特异性的可变亚基[13]。之前使用单粒子冷冻电子显微镜 (cryo-EM) 解析了包含四个Orco亚基的受体的结构[14],阐明了这种同源四聚体通道的基本结构。但是Orco不包含受体的配体结合位点,因此结合位点尚未解析。为了解决昆虫气味受体的完整结构,同时规避解析异多聚体蛋白结构的困难,del Mármol等人将目光集中在一种不需要Orco即能组装成功能性同源蛋白的昆虫气味受体上。
由相似亚基组成的异聚蛋白通常是在演化过程中基因复制时产生的。因此,del Mármol等人推断在演化上更古老的生物体中会存在功能性同聚气味受体,他们重点研究了蠹虫的近亲石蛃Machilis hrabei。在石蛃中,受体库仅由五个亚基 (MhOR1-5) 组成,没有一个与Orco直接相关[15]。每个MhOR5亚基包含九个 α-螺旋,其中六个完全跨越细胞膜(S1-S6),另外两个(S0和S7b)部分跨越细胞膜,最后一个(S7a)位于细胞膜内。
作者测试了由四个MhOR5 或四个MhOR1 亚基组成的通道如何响应不同的配体。他们发现MhOR5 通道被广泛调谐,对他们测试的 60% 以上的化学物质(包括丁香酚和避蚊胺)都有反应,而 MhOR1 通道更具选择性。
接下来作者使用冷冻电镜解析了处于非结合状态(称为 apo 状态)、与避蚊胺、丁香酚结合时的MhOR5通道的结构(图 1)。总体而言,这些结构与Orco同聚体相似。
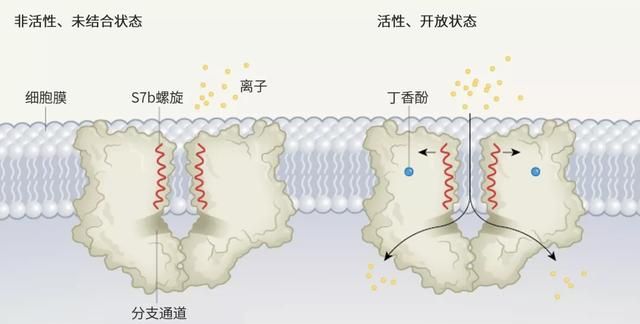

图1 | 昆虫气味受体的闭合和打开状态。
在昆虫中,嗅觉细胞上检测气味分子的受体是由四个蛋白质亚基组成的离子通道。del Mármol 等人[2]使用冷冻电子显微镜解析了来自石蛃(Machilis hrabei)的一种气味受体处于非活性、未结合 (apo) 状态及活性、开放状态的结构。四个相同的亚基构成中心离子渗透通道;这里分别展示了两个亚基。当与丁香酚等气味剂结合时,每个亚基的 S7b 螺旋会轻微旋转,从而扩大细胞膜外表面附近的通道孔。离子穿过一个大的前庭,然后通过四个横向分支通道中的一个进入细胞。
这些结构表明,当通道被激活打开时,带正电的离子沿着一条通道从一个大的外前庭进入单个跨膜的通道,该通道可能是四个亚基中任意一个,均由S7b α-螺旋的氨基酸残基排列而成。在受体的胞内侧,四个通道从中心横向分叉,构成离子进入细胞的通道的胞内部分。在未结合状态下,该通道的最窄部分直径为 5.3 埃,位于 S7b 螺旋中的缬氨酸残基伸入中央管腔的位置,形成疏水性“塞子”,限制离子通过。
未结合和配体结合状态的比较揭示了两个关键的结构特征,这些特征也许能解释气味受体通道通常如何能够对广泛的配体作出反应。首先,通道从关闭状态到打开状态的转变仅涉及 S7b 螺旋的轻微旋转,这会扩大孔径并将极性残基移动到离子通道中。与在电压门控离子通道打开期间 S4 区域的运动相比,这种变化可能只需要很少的能量[16]。通道开启所需能量如此之低,可能是气味受体配体混杂的关键:即使对受体具有低亲和力的配体的结合也会导致重排。
同样重要的是配体结合位点的性质,它似乎异常灵活,能够在多个方面适应不同化学成分的配体。该配体结合口袋位于由 S2、S3、S4 和 S6 螺旋的跨膜部分形成的松散束的深处,内衬主要是大的芳香族和疏水性残基,这些残基与配体形成非极性相互作用。这与在许多其他类型受体的结合位点中观察到的氢键(极性极强)严格的几何限制形成鲜明对比。
此外,MhOR5受体的结合位点似乎会发生重排以适应不同大小和形状的配体。口袋中的细微突变会改变配体特异性,从本质上重新调整受体。这提供了一种机制,即进化可能会产生大量具有不同配体特异性的受体,而这些受体在现存昆虫物种中已经被发现。
终于,在发现脊椎动物气味受体的30年后[6],我们有了气味受体与其配体复合物的结构。但是,尽管这一进展回答了一些古老的问题,它也提出了新的问题。例如,不论是未结合的受体还是配体结合受体的结构都没有揭示气味剂进入配体结合口袋的途径。一种可能性是通道的“呼吸”,暂时打开一条通道,气味剂可以通过该通道到达该部位,从而将通道锁定为开放构象。通过研究受体的结构和功能来定义气味通路可能会对受体特异性有的进一步了解。
这项研究和之前的研究也提出了一个问题,即激活状态下的受体系统在没有配体的情况下(尽管处于低水平)如何区分来自配体结合受体的信号和无配体结合激活的干扰。一种可能性是嗅觉系统可以容忍一定量的干扰,因为它可以随着时间的推移不断收集和整合新信息:例如,当一只昆虫靠近气味源时,它可能会使用新的、更高质量的嗅觉信息来纠正它的路线。另一种可能性是嗅觉系统组织成嗅小球,汇集来自许多神经元的信息,有效地过滤掉单个错误神经元的活动。可能需要分析嗅小球和中枢神经系统对气味信号的处理来回答这些问题。
最后,有人也许还想知道这些发现是否可以推广到脊椎动物G蛋白偶联的气味受体。与昆虫中的气味受体一样,脊椎动物气味受体主要通过非极性和弱极性相互作用与气味结合[17]。它们是否也有灵活的配体结合口袋和低能量屏障来参与下游信号传导,从而使它们能够对各种相对低亲和力的配体做出反应?毫无疑问,del Mármol等人的工作和未来的研究将指引为气味受体设计配体的开发,也许可能有一天会有避蚊胺的替代品。

一个博士的血泪求职路:想入职高校?请以我为鉴避坑
单价日涨3000元,我国6成依赖进口,新能源车无“锂”难行……
Force Dimension 全系列触觉反馈装置:给用户真实的沉浸式体验
特别声明:本文发布仅仅出于传播信息需要,并不代表本公众号观点,侵删;如其他媒体、网站或个人从本公众号转载使用,请向原作者申请,并自负版权等法律责任。
原创文章,作者:芒小种,如若转载,请注明出处:http://www.fhgg.net/shenghuobaike/32944.html
本文来自投稿,不代表【食趣网】立场,如若转载,请注明出处:http://www.fhgg.net/